| ABBREVIATIONS | |
|---|---|
| Abbreviation | Meaning |
| A | anal |
| bsx | basal subcostal crossvein |
| C | costa |
| c1 | first cubital cell |
| c2 | second cubital cell |
| cc | callus cerci |
| Coll. | collecter |
| comb. nov. | combination novum or new combination (Latin novum = new) |
| Cu | cubitas |
| Cu1 | cubitas anterior |
| Cu2 | cubitus posterior |
| dcc | distal cubital cell |
| ed | editor |
| eds | editors |
| ep | ectoproct |
| ent | entoprocessus |
| e.g. | (Latin exempli gratia) for the sake of example. |
| et al | (Latin et alii) and others |
| Fe | femur |
| Fig. | figure |
| Figs. | figures |
| Fl | flagellum |
| Fr | frons |
| ga | gena |
| gl | gonapophysis lateralis |
| gc | gonarcus |
| gcr | gonocristae |
| gm | gram |
| gp | gonopophysis posterior |
| gsc | gonosaccus |
| gst | gonosetae |
| hrs. | hours |
| Ibid | (Latin ibidem) cited just before |
| i.e. | (Latin id est) that is |
| ig | inner gradate |
| im | intramedian cell |
| ml | milliliter |
| km. | kilometers |
| L. | Linnaeus |
| lbp | labial palpus |
| lp | lateroprocessus |
| M | media |
| M1 | anterior media |
| M2 | posterior media |
| M3+4 | fused 3rd and 4th branches of media |
| MA | the proximal branch of fused anterior media and Rs |
| md | mandible |
| mxp | maxillary palp |
| msl | meters from sea level |
| mtsc | metascutum |
| pa | paramere |
| ped | pedicel |
| pp | total number of pages |
| psc | pseudo-cubitus |
| psm | pseudo-media |
| pt | pterostigma |
| R | radius |
| R1 | anterior radius |
| R2 | posterior radius |
| r-m | radio-media |
| Rs | radial sector |
| S | scape |
| Sc | sub costa |
| S.D. | standard deviation |
| Sp | species |
| sp. nov. | species novum or new species |
| sq. km. | square kilometer |
| ti | tibia |
| tar | tarsus |
| viz | (Latin videlicet) namely |
| Vol. | volume |
| SYMBOLS | |
| Symbols | Meaning |
| = | equal to |
| % | percent |
| ºC | temperature in degrees of the Celsius scale |
| ± | plus or minus |
| O | Male |
| ♀ | Female |
The characters by which the Neuroptera are defined are: adults with complete mandibulate mouthparts; two pairs of membranous wings, maybe equal or sub-equal, complex venation, at rest held roof-wise over the back; legs with five tarsal segments, larvae usually carnivorous with developed mandibles and maxillae, pupae exarate.
Different authors have used different terminology in respect of wings, male and female genitalia which are specially important for Neuroptera taxonomy. For structural terminology used for the taxonomy, the book of Imms (1925) [eds. Richards and Davies (1977)] as well as works of Comstock (1918), Tilliyard (1916), Tjeder (1970) and have been followed in the present study. Some of the morphological features relevant to the taxonomic study are given below.
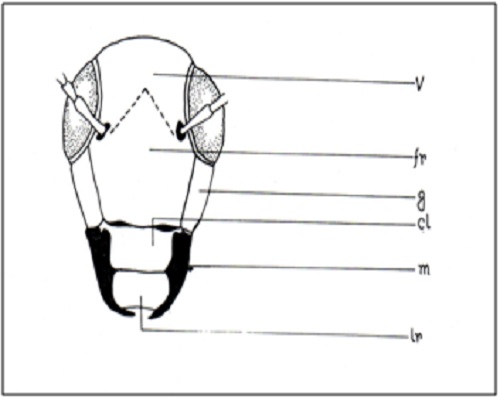
HEAD: (Fig. A, B)Hypognathus with biting and chewing mouthparts in most of the families except in Nemopteridae in which it is protruded in the form of rostrum.
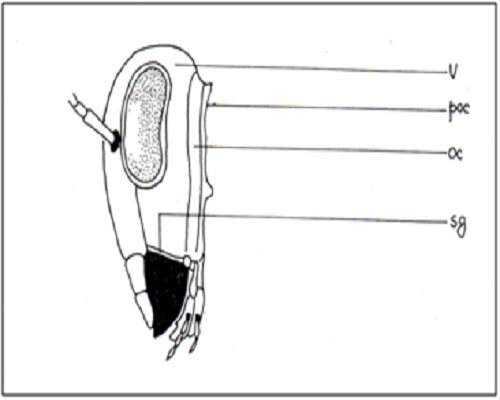
Vertex: Portion of epicranium in between the eyes and top of head, arched dorsally or dome-like but sometimes flattened, unpaired, bears paired ocelli.
Frons: Unpaired upper facial part, separated laterally from gena by frontogenal sulci and distally by epistomal or frontoclypeal sulci, bears median ocelli.
Clypeus: Lies immediately anterior to the frons, divided into large postclypeus and smaller anteclypeus.
Labrum: Unpaired sclerite may be articulated with clypeus by clypeolabral suture.
Occipit: Hinder part of the epicranium between vertex and neck.
Gena: Total lateral area below and posterior to the eye on each side.
Mouthparts: It is formed by a pair of well developed and chitinised mandibles (basal tooth on left mandible may be present); a pair of maxilla, each consisting of cardo, stipes, galea, lacinia and 5 or 6 segmented palpus; a labium consisting of submentum, mentum, prementum, ligula (reduced or absent), and a pair of 3 or 4 segmented palpi.
Compound eyes: Placed laterally on either side of epicranium a pair of large, prominent, widely separated eyes; ocelli three in number when present.
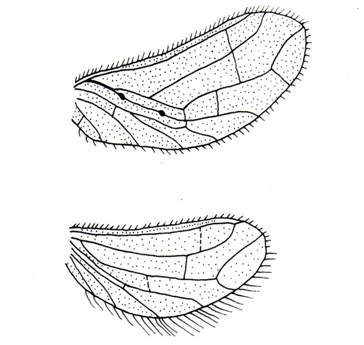
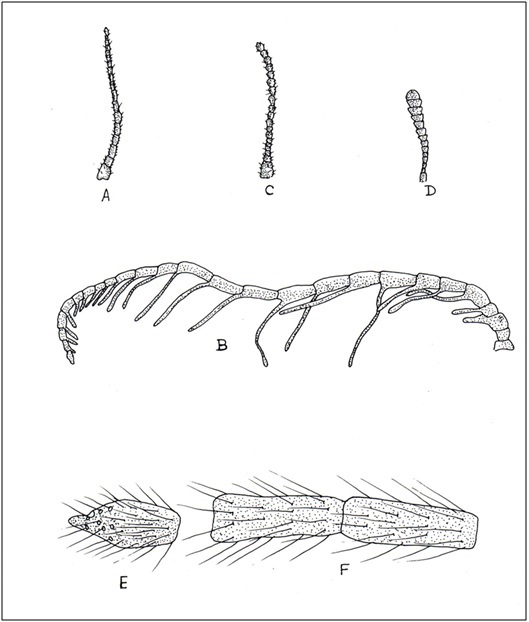
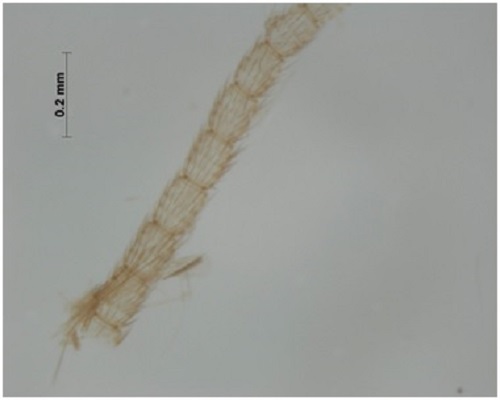
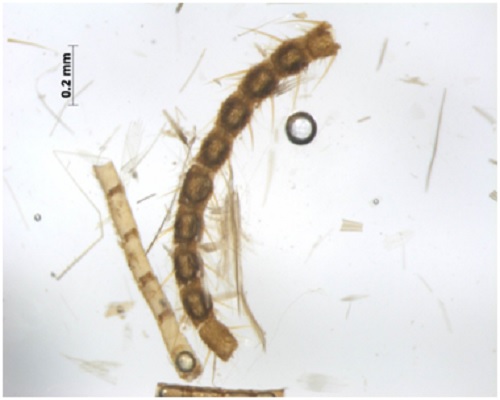
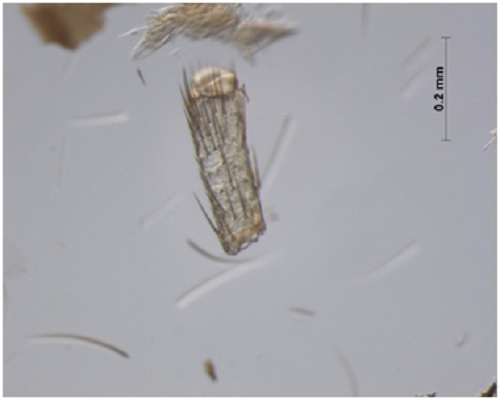
(Fig A-D) Antenna divided into scape, pedicel and flagellum or flagellar segments. Each flagellar segment bears setae which are either arranged regularly in a ring-like formation (Chrysopidae, Hemerobiidae, Coniopterygidae), or in irregular fashion (Dilaridae). Antenna elongated (Hemerobiidae, Chrysopidae, Ascalaphidae), even longer than forewing (Chrysopidae), short (Myrmeleontidae, Psychopsidae), filiform [Fig. A] (Chrysopidae), monaliform [Fig. C] (Hemerobiidae, Berothidae), pectinate [Fig. B, Plate III] (male Dilaridae), clavate [Fig. D] (Ascalaphidae), or capitate (Myrmeleontidae).


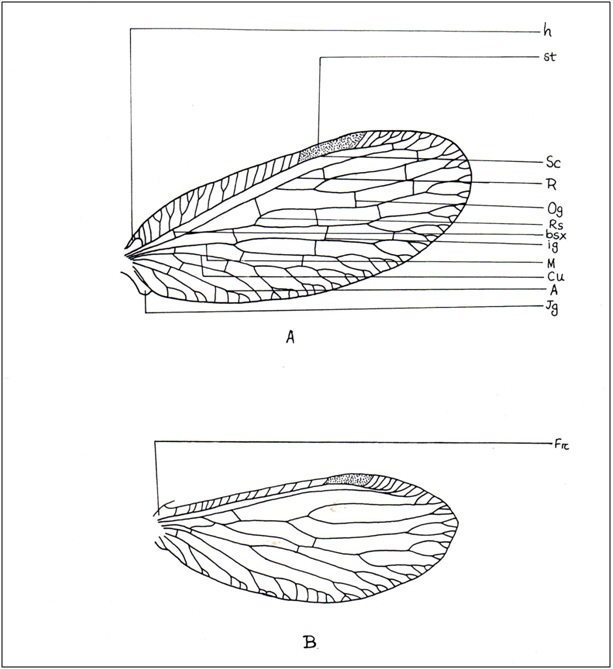
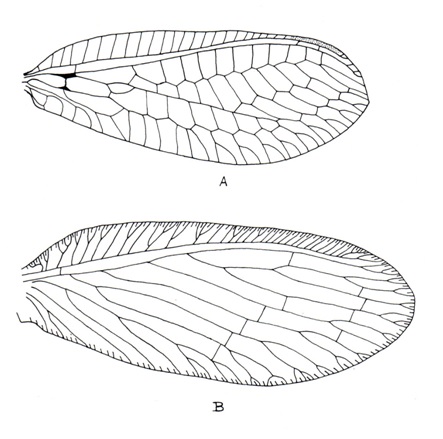
(Fig. A-B) Two pairs; usually subequal (but hind wing ribbon, racket or thread-like in Nemopteridae); generally oval or elongated, at rest held vertically over dorsum; membranous, membrane usually delicate and smooth; may be thickened and coriaceous in some species; generally hyaline, sometimes variably clouded or eroded by powdery secretion (Coniopterygidae) or veins under cover of seed like scales (females of Berothidae). Hindwing may be reduced or absent (Hemerobiidae). Trichosors always present (Hemerobiidae, Dilaridae, Psychopsidae) or absent (Chrysopidae); may be fused with vein-ends forming a list-like margin (genus MicromusRambur, Hemerobiidae); crossveins always with microtrichia or macrotrichia or without, in one Micromus species extremely developed into long spine. Pterostigma generally distinct (Hemerobiidae, Chrysopidae, Myrmeleontidae) or absent (Dilaridae).Costal area narrow (genus MicromusRambur, Hemerobiidae) or enormously broad (Psychopsidae). Generally a wing coupling apparatus is present but maybe absent (Hemerobiidae). A triangular jugal lobe is found at base of anal margin. Basally hind wing carry a distinct frenulum.
The characteristic Neuropteran venations are as follows:
C: Costal area narrow or broad, costal veinlet numerous or absent (Coniopterygidae), may be interconnected by crossveins (Psychopsidae) or not (Chrysopidae, Hemerobiidae); humeral vein absent or rudimentary (Berothidae) or present (Hemerobiidae), forked or unforked (Chrysopidae).
Sc: A strong vein meeting C near apex.
R: A strong vein divided into R1 and Rs.
R1: Long, parallel to Sc, running separate or fused with Sc (Myrmeleontidae, Ascalaphidae) creating hypostigmaticcell (which is long in Myrmeleontidae, short in Ascalaphidae), connected with Sc by crossvein or not (Dilaridae).
bsx: Connecting Sc and R at base.
Rs: One with numerous crossveins (Chrysopidae) in between R1 and Rs or only 3 main branches with 1-2 crossveins between R1 and Rs (Dilaridae) or more than one sector (Hemerobiidae, Coniopterygidae). In PsychopsidaeSc, R and Rs strong and closely parallel forming 'vena-triplica' supported by several crossveins.
psc: Formed by Cu1, distal portion of M 1+2, M 3+4 and by part of three proximal branches of Rs, distinct in Chrysopidae.
psm: Formed by crossveins of Rs and M.
c1, c2, dcc: These cells formed in between Cu1 and Cu2. In Berothidae, Cu1 of hind wing runs for a long distance close to and generally parallel to the hind margin, Cu2 may be indistinct or absent.
1A, 2A, 3A: Usually present. 1A forked in genus Nothochrysa McLachlan (Chrysopidae).




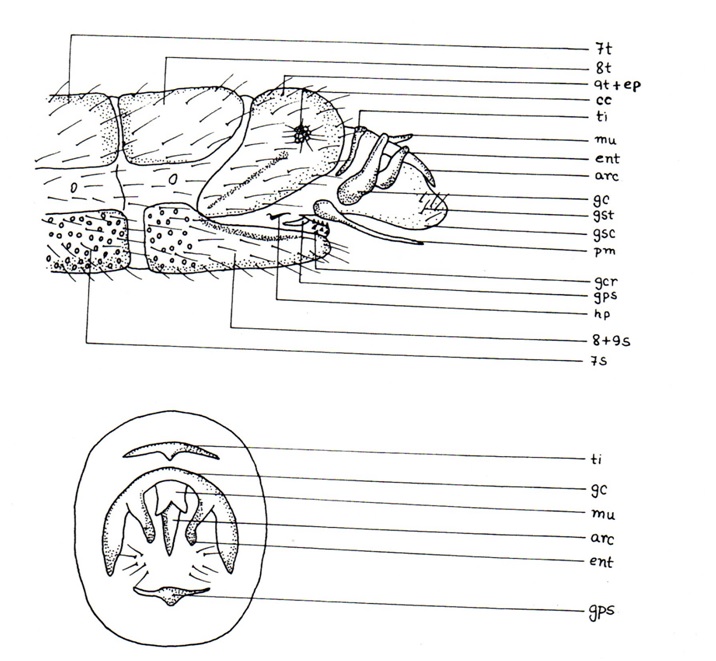
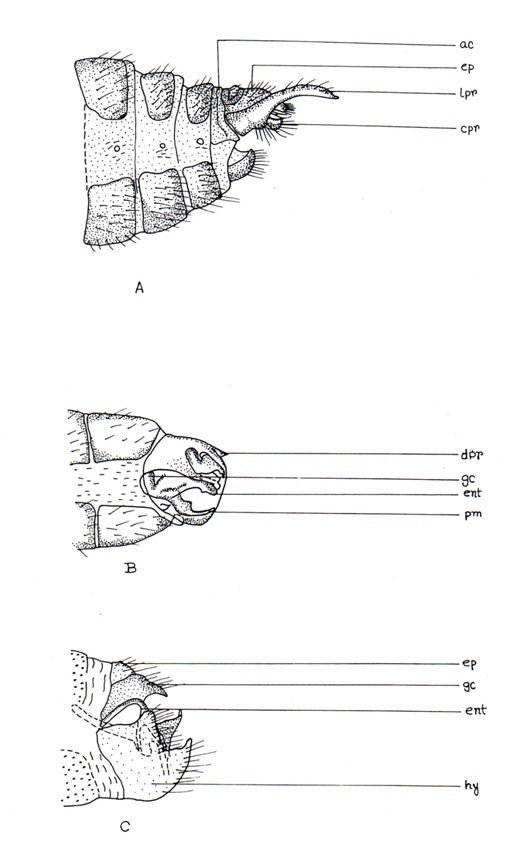
The 8th segment discleritous and usually separate from 9th which is either synscleritous or discleritous, but in most Chrysopidae 8th and 9th sternites are fused. 9th tergite is either synscleritous or discleritous, mid-dorsally divided into a pair of plates, 9th sternite in majority is a simple plate or half-ring or an elongated one, covered from above by the lower part of the membrane that form the hind body wall of the abdominal end. In many members of the Chrysopidae, dorsal surface of these sternite membrane has a single or a pair of sclerotized plates, carrying strong teeth called gonocristae. In some species of Chrysopidae, an usually arched structure internally fused to the membrane and gonocristae. The free end of this structure may be simple or forked and extending through the membrane. This structure is called gonapsis. Small, dome-like hollowed structures between the hairs, themicrotholi, present on surface of some Chrysopidae in 8th and 9th segments. The 9th segment may have a backwardly directed projection on each side, called lateroprocessus, present in some Hemerobiidae (Fig. 6A) and a single median backwardly directed process, the dorsoprocessus, present in Dilaridae (Fig. 6B). A strongly sclerotized bar, the antecosta, may sometimes be attached or separated along the proximal margin of the 9th tergite. Inside the 9th tergite and sternite bar-like apodemes frequently occur, which in some Chrysopidae project beyond the apex of the sternite and develop into a very coarse structure, the solimeres. In Raphidiidae, Inocelliidae and Coniopterygidae a pair of gonocoxites are often associated with the 9th sternite, but in Coniopterygidae they may fuse with each other to form external hypandrium (Fig. 6C), with a median apical incision forming two processusterminalis, one on each margin of hypadrium. Hypandrium may be telescopically arranged in the 9th sternite of Aleuropteryginae.
The phallic structures are situated between the anus and the 9th sternite. A sclerotized intromittent organ or penis present only in Coniopterygidae. Other families have an elastic opening, gonopore, on the hind body wall membrane which form an inverted, sometimes paired sac, the gonosaccus, with a number of strong hairs, gonosetae, or with a number of minute teeth, spinellae. In Nemopteridae, a sclerotized, dark pigmented area, gonolatus, is present. Ascalaphids have finger-like projections, pulvilli, from gonosaccus, whereas an unpaired, long, tubular organ, pseudopenis, is found in some Chrysopidae. The predominant structure of the phallic complex is the gonarcus, present in all families, situated dorso-basally, generally an arch-shaped structure with its arches directed downwards or inwards. The gonarcus may be dorsally divided into a pair of plates, when undivided its median parts ending into a backwardly directed tooth-like process, mediuncus or median plate. A forked, unpaired immovable, downwardly directed process, hypocuspis present under mediuncus in Nymphes. Each gonarcus arch may have a lateral process, entoprocessus. In many genera, an additional vertically movable structure, arcessus, is present below the central part of gonarcus. Another structure, movably attached, hypostylus, may be present in the lowest part of gonarcus (Nymphes). In Osmylidae an additional narrow structure, buculum, is present on each side of gonarcus. Below the gonarcus and fused with it, a pair of parameres are present. They are frequenly fused with one another, when fused at the proximal end they form apophysisproxima (Hemerobiidae). Apophysislateralis is also present when the parameres fuse laterally. Generally each paramere has a dorsal backwardly directed supraprocessus (Hemerobiidae). An apical, upwardly directed structure, processusapicalis, is connected with the main stem of parameres in Coniopterygidae. A shield-like, ovate or elongated plate with sensory hairs, pelta, is often hidden in between the distal part of the parameres of Ascalaphidae and Myrmeleontidae. Ventral to the phallic structures, downwardly projected, is a pair of hypomeres found in Berothidae and Mantispidae. In some Chrysopidae a similar arch-formed structure, the tignum, is present dorsal to the gonarcus. In Belonopterygini (Chrysopidae) the gonarcus is almost transverse often with a pair of lateral horns, the gonocornua.
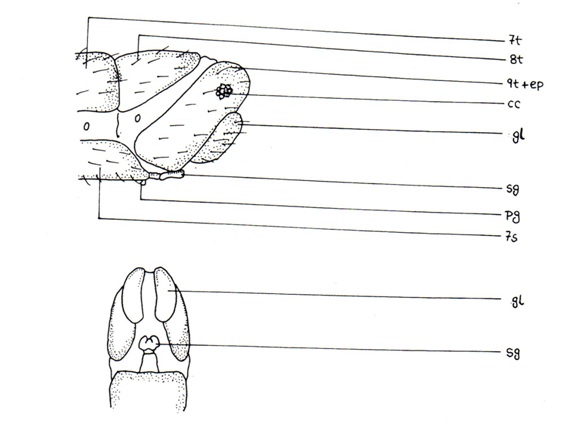
A peculiar stem-like organ, the hypandriuminternum, present in all families, is situated at the base of ductusejaculatorius just at the place where the two gonoduct unite. Proximally situated and membranously attached to the hypandriuminternum in some Chrysopidae, there is an organ, comes. The annal segments (anoprocessus, catoprocessus and cercus present individually in primitive Corydelidae) are fused together to form a single lateral plate, the ectoproct. The cercus is reduced to callus cerci bearing trichobothria or is totally absent. In Hemerobiidae, two ectoproct are free for grasping, each may have one prong or two, representing either anoprocessus or catoprocessus or both. Below the anus, a weakly sclerotized hairy plate, the subanale, is present (Chrysopidae). In some Dilaridae, the ectoproct is absent, a strongly sclerotized supra-anale and a less sclerotized proctiger hold the anus.
The 8th segment frequently consists of a dorsal half-ring. This often has downard prolonged sides, sometimes fused ventrally with a secondarily developed sternite (Hemerobiidae). 8th sternite generally missing. Instead of 8th sternite a subgenital plate is present, subgenitale, bilobed in Chrysopidae, maybe absent in Hemerobiidae. Another structure, praegenitale, is present with distal lower apex of 7th sternite, behind subgenitale, it is rare in Hemerobiidae. Long, finger like, backwardly projected lateral appendages, gonapophysisanteriores present in Myrmeleontidae.
The 9th tergite forms a half ring or divided longitudinally or laterally, sometimes fused with the ectoproct. In Sisyridae and Ithonidae the tergite halves are almost sternite like. When it is laterally divided, creates dorsal plates and lateral plates. Lower portion of 9th tergite may have a finger like projection, lateroprocessus. A secondary 9th sternite is developed in some Coniopterygidae. Lateral side of ovipositor covered by gonapophysislaterales, membranously connected dorsally and have a longitudinal ridge like apodeme. Some Hemerobiidae and Nemopteridae have a finger like slender organ under gonapophyseslaterales, the hypocauda. Gonopophysesposteriores present above the apex of subgenitale, inserted inside gonopophyseslaterales.
Lower part of the 9th segment with gonapophyseslaterales form the genital chamber in which common oviduct and bursa copulatrix open. Bursa copulatrix is usually membranous (sclerotized and pigmented in Chrysopidae, Coniopterygidae and Sialidae), spermatheca strongly sclerotized and pigmented (weak in Hemerobiidae) with variable shape. It is a long tube in Myrmeleontidae; coiled duct in Berothidae; twisted sac in Hemerobiidae; a round, flattened box with dorsal triangular process, the vela, and a ventral impression in Chrysopidae; maybe a paired vesicle in Raphidiidae or a single glandulaaccessoria.
An ectoproct, situated laterally on each side of the anus or the two ectoprocts are fused above the anus or even with the 9th tergite. Callus cerci absent or present with trichobothria.\